Immune System Series
Receptors for Recognizing Antigen
antigenIn order to recognize and respond to the antigens that are their specific targets, both B cells and T cells carry special receptor molecules on their surface. For the B cell, this receptor is a prototype of the antibody the B cell is prepared to manufacture, anchored in its surface. When a B cell encounters a matching antigen in the blood or other body fluid, this antibody-like receptor allows the B cell to interact with it very efficiently.
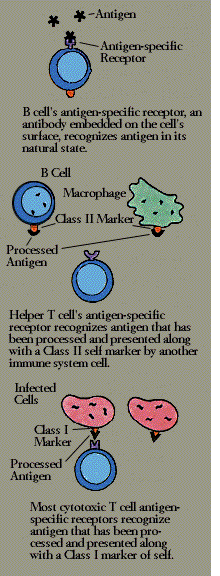
The T cell receptor is more complex. Structurally, it is somewhat similar to an antibody, made of a pair of chemically linked chains with variable and constant regions. (But to work, it needs the help of an associated set of signaling and anchoring cell surface molecules called T3.) Unlike a B cell, however, a T cell cannot recognize antigen in its natural state; the antigen must first be broken down, and the fragments bound to an MHC molecule, by an antigen-presenting cell.
Helper T cells (T4 cells) look for antigen bound to a class II MHC molecule-a combination displayed by macrophages and B cells. Most cytotoxic T cells (T8), on the other hand, respond to antigen bound to MHC class I molecules, which are found on almost all body cells.
The T cell receptor molecule thus forms a three-way complex with its specific foreign antigen and an MHC protein. This complicated arrangement assures that T cells-which affect other cells through either direct contact or bursts of secretions-act only on precise targets and at close range.
The major antigen receptor, name alpha/beta for its two chains, is found on most T4 and T8 cells. A second, more recently discovered antigen receptor also has two chains and is known as gamma/delta; it is found on a distinct subset of mature T cells. Like the alpha/beta receptor, the more primitive gamma/delta receptor works in conjunction with T3. The function of T cells that carry gamma/delta receptors is not known.
Humoral immunity chiefly involves B cells, although the cooperation of helper T cells is almost always necessary. B cells, like macrophages, take in and process circulating antigen. Unlike macrophages, however a B cell can bind only that antigen that specifically fits its antibody-like receptor.
To enlist the help of a T cell, the B cell exhibits antigen fragments bound to its class II MHC molecules. This display attracts mature helper T cells (which may have been already activated by macrophages presenting the same antigen). The B cell and T cell interact, and the helper T cell secretes several lymphokines. These lymphokines set the B cell to multiplying, and soon there is a clone of identical B cells. The B cells differentiate into plasma cells and begin producing vast quantities of identical antigen-specific antibodies.
Released into the bloodstream, the antibodies lock onto matching antigens. The antigen-antibody complexes trigger the complement cascade or are removed from the circulation by clearing mechanisms in the liver and the spleen. The infection is overcome and, in response to suppressor influences wielded by yet other subsets of T cells, antibody production wanes.
Clinically, infections manifest themselves through the five classic symptoms of the inflammatory response-redness, warmth, swelling, pain, and loss of function. Redness and warmth develop when, under the influence of lymphokines and complement components, small blood vessels in the vicinity of the infection become dilated and carry more blood. Swelling results when the vessels, made leaky by yet other immune secretions, allow fluid and soluble immune substances to seep into the surrounding tissue, and immune cells to converge on the site.